From Advances in New Crops,
Proceedings of the First National Symposium NEW CROPS: Research,
Development, Economics
by D. N. Moriconi, M. C. Rush, and H. Flores
Tomatillo: A Potential
Vegetable Crop for Louisiana
1. INTRODUCTION
2. BOTANY
1. Plant Characteristics
2. Production
3. HORTICULTURE
4. TISSUE CULTURE
5. FUTURE PROSPECTS
6. REFERENCES
7. Fig. 1
8. Fig. 2
9. Fig. 3
10. Fig. 4
INTRODUCTION
Throughout
history humans have used some 3000 plant species for food. The recent
tendency has been to exploit fewer and fewer species and today, only
around 20 species supply most of the world's food. Many beneficial
plant species have been underused or have not been developed to their
full potential (Vietmeyer 1986). Useful plant species have often been
overlooked because they are native to the tropics, regions neglected by
the world's research institutions which are oriented toward crop
production in temperate zones.
There are several Solanaceous species with edible fruit that are
popular in Latin America in addition to tomato and chilies. Physalis ixocarpa
Brot.; commonly known as the husk tomato and by the Spanish names of
tomate de cascara, tomate verde, tomate de fresadilla, tomatillo, and
miltomate, is an important vegetable crop in the diets of Mexicans and
Central Americans. In Mexico, the fruits are used in the making of
chili sauce and dressings for popular dishes such as tacos and
enchiladas. P. ixocarpa
is
gaining ground as a new crop in California due to the increased
popularity of Mexican food in the United States (Quiros 1984) and has
production potential in the southern United States. In Louisiana,
tomatillo imported from Mexico is sold as a fresh fruit in a few
grocery stores. There is a potential market for fresh produce and the
Louisiana sauce industry may be interested in opening a new ethnic
market for their products. Developing a new crop is a difficult and
complex process. One way is to import an exotic crop from areas where
the crop is already grown or consumed and adapt it to local conditions
(Laidig et al. 1983). It appears that P. ixocarpa
has potential as a commercial crop in Louisiana. Our interest in
tomatillo is to satisfy the regional demand for fresh product and to
develop a new processed product for the Louisiana canning and sauce
industry.
A variety of world-wide, national and local economic
factors have combined to cause a reduction in the income of Louisiana
farmers. Along with other areas of the country, Louisiana has begun to
search for crops that would diversify its agriculture (Hamm 1985). The
estimated net return per unit area of land is high in vegetables when
the volume of production and sales is sufficient to amortize the
required investment Hinson and Cannon (1988) consider that 350,000 to
400,000 ha are suitable for vegetable production in Louisiana. The
light-textured alluvial soils along the Mississippi, Red, and Ouachita
rivers are highly appropriate for vegetable crops. Climate and soil
type are major factors that must be considered for vegetable
production. Generally, the rainfall in Louisiana is sufficient for crop
production without irrigation; however, the pattern of rain is such
that some crops could be destroyed by excessive rainfall unless
drainage is provided. In other areas, irrigation after transplanting
would be necessary for survival of transplants. The area planted to
vegetable crops has increased from 4,300 ha in 1980 to 7,300 ha in
1986. Louisiana clearly has the resources for producing vegetable
crops. Both land and water are abundant, although water must be
managed. A local supply of labor is available, due to the high
unemployment rate (Hinson and Cannon 1988).
Louisiana has a
total of 15 commercial fruit and vegetable processing plants. Six of
the plants are exclusively canning factories. Eight plants produce
pickled products, hot pepper sauce, and other table sauces for
seasoning. The remaining processing plants produce products in bulk for
further processing. Except for two plants, the processing plants in
Louisiana are located in the south central section. Sauce plants are in
Saint Martin parish (Acadian Pepper Co., Bruces Food Co., Cajun Chef
Products Inc., and Landry Brothers), Iberia parish (Durke-French Foods
Inc., McIlhenny Company East, and B.F. Trappey's Sons, Inc.), East
Carroll parish (Panola Pepper Co.), and Orleans parish (Baumer Foods
Inc.) (Broussard and Hinson 1988).
The total volume of table
sauces, pickled, and other items processed in Louisiana is around
22,277,000 kg with an estimated value of $58,427,000. Table sauces
accounted for approximately 77% of the total volume (Broussard and
Hinson 1988). The increase in table sauce production may be a result of
the popularity of cajun food observed across the U.S. Studies have been
conducted toward developing tomatoes for processing in Louisiana, but
the high summer temperatures cause flower abscision and reduce crop
yield. Tomatillo plants are known to grow and set fruit at high
temperatures. The Yucatan peninsula, the main area for tomatillo
production in Mexico, has daily mean temperatures similar to those in
Louisiana during the summers and maximum temperatures are higher in the
Yucatan throughout the year than in Louisiana (Fig.
1).
The increase in the popularity of Mexican food in the last decade, and
the amount of Mexican sauce available in the grocery stores indicates a
high potential for tomatillo as an ingredient of canned taco and
enchilada sauces. Tomatillo, a main component in traditionally made
taco and enchilada sauces in Mexico and Central America could be
produced in Louisiana and sold in the fresh vegetable markets of the
Southern U.S. or processed by Louisiana companies in sauces.
BOTANY
The genus Physalis,
established by Linneaus in 1753, contains about 100 species of annual
and perennial herbs (Willis 1966). The genus is characterized by the
presence of pendant flowers and an inflated fruiting calyx which
encloses the berry (Sullivan 1984). Four species are cultivated in
different parts of the world for their fruit: P. peruviana L.
(cape gooseberry, uchuba) and P.
pruinosa L. (ground cherry, husk tomato) are used as jam
fruits; P. alkekengi
L. (Chinese lantern) is used as an ornamental; and P. ixocarpa Brot.
(tomatillo, tomate de cascara) is used as a vegetable or for sauces.
Several species of Physalis
are widespread in America as endemic weed species. Six important Physalis
spp. are prevalent in the phytogeographic region of Mesoamerica
(Belize, Guatemala, Honduras, El Salvador, Nicaragua, Costa Rica, and
Panama, and the Mexican states of Chiapas, Yucatan, and Quintana Roo): P. angulata L., P. cordata Mill., P. gracilis Miers, P. ignota Britt., P. lagascae R.
& S., and P.
pubescens L. (Gentry and D'Arcy 1986). These Physalis
spp. can be intercrossed, but incompatibility has been found (Pandey
1957, Quiros 1984). The basic chromosome number of the genus is N=12
and most species are diploid; P.
peruviana is a tetraploid (Menzel 1951).
Tomatillo has been known to botanists for nearly 400 years as P. philadelphica
Lam. Francisco Hernandez in 1651 described two varieties from numerous
plant types called tomate by the Aztecs. Botanists have suggested that
the small-fruited miltomate is a wild-type plant, whereas, the
tomatillo is a domesticated plant that derives from plants similar, if
not identical, to miltomate (Hudson 1986). The specific boundaries in Physalis
are poorly defined with some duplication of names and many changes in
the nomenclature during the last 50 years. The complexity of the genus
is caused mainly by the wide range of genetic variability present
presumably resulting from interspecific hybridization (Menzel 1951,
1957; Waterfall 1958) and also by the ambiguity of the earlier
taxonomic descriptions (Raja-Rao 1979). For example, P. aequata Jacq.
and P. capscicifolia
Rydb are considered synonymous with P. ixocarpa.
To clarify the taxonomic classification of Physalis, Menzel
(1951, 1957) and Waterfall (1967) made extensive cytologic and
taxonomic studies of the genus. Menzel reduced P. philadelphica to
synonymy under the variable P.
ixocarpa
Brot. a name that had to come to be widely used for the domesticated
tomatillo (Hudson 1986). The only apparent difference between the two
species was the length of the peduncle, with the peduncle of P. ixocarpa shorter
than that of P.
philadelphica.
Waterfall (1958) accepted this nomenclature when studying the species
of North Mexico, but he reversed himself when he analyzed Physalis spp. from
Mexico and Central America (Waterfall 1967). He incorporated the
small-flowered P.
ixocarpa within the broader limits of P. philadelphica.
Fernandes (1974) made a thorough investigation of this nomenclatural
problem and concluded that P.
ixocarpa is a distinct species, different from P. philadelphica
based on previous cytological evidence, the distinctive sigma, and the
small flowers of the type. Chromosome morphology has recently been used
to understand the interspecific relationships in the genus. Gottschalk
(1954), Raja-Rao (1979), Venkateswarlu and Raja-Rao (1977, 1979a, b),
and Raja-Rao and Lydia-Prasad (1984) studied the morphology of
chromosomes during the pachytene stage with most important Physalis
spp. and demonstrated cytological differences between the species.
Nevertheless, the taxonomic complexity of the genus is not yet
clarified, especially between P.
ixocarpa and P.
philadelphica.
HORTICULTURE
Plant
Characteristics
Plants in the genus Physalis
have herbaceous stems. Some have short to elongated rhizomes, the
leaves are usually broadly ovate to linear and generally alternate. The
flowers are solitary in the axis of the leaves, sometime pendant in the
axillary branches causing them to appear to be axillary between the two
branches. The pendant blossoms are often hidden by the foliage and many
of the flowers hang just above the ground (Sullivan 1986). The flowers
have corollas campanulate to rotate with the petal borders reflexed.
Petals are usually yellow with a dark purple spot near the base of each
petal. The calyx is united, with lobes more than one half its length.
The androecium has five stamens with the filaments attached to the base
of the corolla tube. The anthers are ovate-oblong and dehiscent by
lateral slits. The fruit is a two carpet, many seeded-berry (Waterfall
1958). There are several reports concerned with the development and
growth of tomatillo plants (Mulato-Brito et al. 1985; Cartujano-Escobar
et al. 1985a, b), and we have 2 years of experience with tomatillo
growing in Louisiana.
Tomatillo seedlings form a single shoot
which has three to five internodes above the cotyledons. The last
internode ends with a flower, one leaf and two lateral ramifications.
Each ramification has one node which terminates in the same pattern,
one end flower, one leaf and two branches. This pattern continues until
senescence, "with the exception that when two leaves are formed there
is no further branching (Fig. 2).
One
characteristic of the main branches is that the internodes differ in
length and have many adventitious roots. When these roots contact soil,
they grow into the soil and are independent of the main root system.
The
number of fruits set is variable, but generally fruit are set until the
10th and 11th weeks after emergence. Tomatillo is similar to tomato
plants, in that the biggest fruit are from the first flowers on the
main branches. The lateral and sublateral branches produce more flower
buds but they abscise and do not produce harvestable fruit.
Mulato-Brito et al. (1985) reported that the greater total number of
nodes on the lateral and sublateral branches produce more fruits than
the main branches, but that those fruits rarely reach commercial size.
Maximum fruit production is reached by 11 weeks after emergence. The
high number of fruits present at this time compete with each other for
the available nutrient supply. Most of the fruit on lateral branches
are
dropped or they do not reach commercial size (Cartujano-Escobar et al.
1985b).
A breeding program should select for a determinant plant
type suitable for mechanical harvesting of tomatillo. The elimination
of sublateral branch production and reduction of internode number in
lateral branches would be very important for restricting fruit
production to a short period of time.
The seed germinates in
7-10 days, followed by elongation of the primary shoot for 4 weeks. The
first flower bud is formed before the elongation of the primary shoot
end, which is around the 3rd or 4th week after emergence, and flowering
continues until senescence of the plant. The first flower appears 4 to
5 weeks after emergence and the first fruit appears one week later,
reaching 3 cm in diameter at 8 weeks. The symptoms of senescence are
visible after 12 weeks, with the plant reaching total senescence at 14
or 15 weeks after emergence (Fig. 3).
The
growing period for tomatillo is short (3 to 4 months) and several
overlapping crops could be produced in Louisiana. The only limit to
plant growth is low temperatures. The growth of tomatillo is poor at
temperatures of 16-18°.C or less. Plants grown in Louisiana
during the
hottest part of the summer produced marketable size fruits (up to 7 cm
in diameter).
Production
Tomatillo
is an important vegetable in Mexico and Central America, where it is
claimed that there is no acceptable substitute for this fruit in making
green sauce (moles) which is served together with regional dishes
(Saray-Meza et al. 1978). Its consumption in central Mexico is about
10% of the total consumption of tomato (Cartujano-Escobar et al.
1985a). In Mexico and Guatemala it is common to find escaped tomatillo
as a weed. Tomatillo originated in Mexico and probably was domesticated
in pre-Columbian times. The plant is an annual, 1-1.5 m in height,
acclimatized to tropical-subtropical humid conditions. These plants
have numerous branches in a dichotomous pattern. The fruit is enclosed
in a husk (enlarged purple veined calyx), but unlike the cape
gooseberry, the inflated calyx stops growing before the berry and is
usually split by the expanding berry (Fig.
4).
The berry is large, round, sticky, green or purplish, high in ascorbic
acid (36 mg/100 g), nicotinic acid (3.5 mg/100 g) and in solids (9%) as
compared to the tomato (6%) (Yamaguchi 1983). The pulp is glutinous, a
little sweeter than tomato, and the flavor is somewhat similar to apple
according to Herklots (1972). The fruit is normally cooked before it is
consumed.
Mexico.
Tomatillo is cultivated in Mexico from the second week of May to the
middle of December, which would correspond to temperatures from mid-May
to mid-October in Louisiana. It is cultivated with and without
irrigation. Tomatillo in central Mexico follows the culture of
sugarcane (Saccharum
officinarum L.) or rice (Oryza sativa
L.). These are also major crops in Louisiana. Thus, the crop rotation
and climatic conditions would be similar in Louisiana. Tomatillo
requires a well prepared soil, generally with furrows 25-cm deep, and
intensive tillage to allow good development of the root system
(Grazon-Tizanado and Garay-Alvarez 1978).
The number of harvests
in Mexico varies with the plant type and quality of the product. Four
to six harvests is normal. Fruit removal should begin when three or
four fruit are mature on each plant which is around 55 to 70 days after
transplanting. A fruit is considered mature when the berry fills the
husk and in some cases breaks it. The size of husk and fruit, the
color, and the flavor of the fruit is variable. The fruit can be green
to yellowish-green or even purple and the flavor can range from sweet
to acid sweet (Villanueva and Loya-Ramirez 1976). The criollas types of
tomatillo yield about 15,000 kg/ha in Mexico. The cultivar `Rendidora'
yields about 25,000 kg/ha. `Rendidora' had about 35% of the total
production in Mexico with large (5 to 7 cm) size fruit and 85% of the
fruit were of commercial quality (Saray-Meza et al. 1978).
Tomatillo
is self-incompatible, so all plants are hybrids. Pollination is by
insects. Cross pollination with other cultivars or other Physalis
spp. would be possible if the plants are closer than 500 m. All seed
production must be carried out in isolation. Saray-Meza et al. (1978)
reported that 10 kg of fruit yields 100 to 200 g of seeds. Plant
viruses can reduce tomatillo yields by 30 to 40%. Delgado-Sanchez
(1986) described a complex of at least three different viruses
affecting tomatillo.
Louisiana.
Plants of P. ixocarpa
were grown in the greenhouse in 1986 with seeds from a single fruit.
Seeds were germinated in petri plates with wet filter paper. The
plantlets were transferred to 7.5 cm pots and placed in the greenhouse.
When plants reached 4 or 5 leaves (4 weeks), they were transplanted to
the field. The field was ploughed twice at 25-30 cm deep, fertilized
with 50 kg/ha 15-15-15 (NPK) and covered with black plastic mulch
before transplanting. Rows were 120 cm apart with 60 cm between plants.
Tomatillo plants were transplanted to the field on June 6, June 25,
July 15, and August 1. Insecticide was applied at 15 day intervals. The
first harvest was made after 6 weeks and harvesting continued at 10 day
intervals for a total of seven harvests during the plant cycle. The
estimated yield was 13,450 kg/ha. There was variation between plants in
size, leaf shape, fruit size and shape, and yield. Fruit damage by
lepidopterous insects was severe, probably reducing the yield by 20 to
30%. No major diseases were observed.
TISSUE CULTURE
A
tissue culture system to exploit somaclonal variation in tomatillo was
developed in our laboratory (Moriconi et al. 1988). Leaf disc or
hypocotyle and epicotyle explants were plated onto Murashige-Skoog
medium (MS) (Murashige and Skoog 1962) containing 3% sucrose, 0.25%
Gelrite or 0.8% agar, B5 vitamins (Gamborg et al. 1968) and 4 mg/liter
2,4-dichlorophenoxyacetic acid (2,4-D). Callus forming on the explant
pieces was transferred to basal MS medium supplemented with 1.0
mg/liter benzylamino-purine (BA) and 0.5 mg/liter indole-3-acetic acid
(IAA). Shoots from organogenesis or embryogenesis formed on the callus
after 6 to 8 weeks of incubation at 26°C, under cool white
fluorescent
lamps, using a 16 hour photoperiod. These shoots developed into normal
plants and could be transferred to a peat-soil mix in the greenhouse.
For
micropropagation, single nodes from sterile regenerated plants were
subcultured in Magenta GA7 vessels containing a medium consisting of
basal MS, B5 vitamins, 0.25% Gelrite, and 10 g/liter sucrose. These
plated nodes readily proliferated shoots from buds. Surface sterilized
stem pieces from seed-grown plants also produced numerous shoots from
buds when plated on this medium. Micropropagation will be useful in
maintaining lines for hybridization for seed production and for
producing large numbers of cloned plants from high yielding,
horticulturally superior plants.
FUTURE PROSPECTS
The
prospects for utilizing tomatillo in the sauce industry are excellent
Sauce made from tomatillo by the Herdex company in Mexico, is being
distributed in the United States by Festin Food Corporation of
Carlsbad, California. Tomatillo is also the main constituent of the
taco sauce packed and distributed by La Victoria Foods, Inc. of City of
Industry, California. Presently Mexico is the source of tomatillos used
commercially in the United States. Local sources of tomatillo should
find an industrial market if a consistent supply can be provided
economically.
Tomatillo is genetically highly variable. To
become a viable commercial crop it will be necessary to develop plants
with uniform fruit size suitable for mechanical harvesting. Mechanical
harvesting requires a determinate plant with most of the fruit maturing
at about the same time. The husk should be loose at maturity and the
fruit should detach easily from the pedicel. Breeding programs should
give these characteristics priority
REFERENCES
Broussard, K.A., and R.A. Hinson. 1988. Commercial fruit and vegetable
processing operations in Louisiana, 1986-1987 season. Louisiana Agr.
Exp. Sta. A.E.A. Info Ser. 68. Louisiana State University, Baton Rouge.
Cartujano-Escobar, F., L. Jankiewicz, V.M. Fernandez-Orduna, and J.
Mulato-Brito. 1985a. The development of the husk tomato plant (Physalis ixocarpa
Brot) I. Aerial vegetative parts. Acta Soc. Bot Pol. 54:327-338.
Cartujano-Escobar, F., L. Jankiewicz, V.M. Fernandez-Orduna, and J.
Mulato-Brito. 1985b. The development of the husk tomato plant (Physalis ixocarpa
Brot) 11. Reproductive parts. Acta Soc. Bot. Pol. 54:339-349.
Fernandes, R.B. 1974. Sur
I'identification d'une espece de Physalis
souspontanee au Portugal. Bol Soc. Brot. 44:343-366.
Gamborg, O.L., R A. Miller, and K. Ojima. 1968. Nutrient requirements
of suspension cultures of soybean root cells. Exp. Cell Res. 50:151-158.
Gentry, J.J., and W.G. D'Arcy. 1986. Solanaceae of Mesoamerica. p.
2-26. In: W.G. D'Arcy (ed.). Solanaceae, biology and systematics.
Columbia Univ. Press, New York.
Gottschalk, W.
1954. Die chromosomenstruktur der solanaceae unter berucksichtigung
phylogenetischer fragestellungen. Chromosoma 6:539-626.
Grazon-Tiznado, J.A., and R. Garay-Alvarez. 1978. El cultivo del tomate
de cascara en el estado de Hidalgo. Circular CIAMEC N 58. Mexico.
Hamm, S.R. 1985. Profile: consumption and production of the U.S.
vegetable industry p. 4-13. In: E. Estes (ed.). Proc. of analyzing the
potential for alternative fruit and vegetable crop production seminar.
North Carolina Agr. Res. Serv- and Tennessee Valley Authority
Herklots, G.A. C. 1972. Vegetables in
South-East Asia. Hafner Press. New York p. 372-376.
Hinson, R.A., and J.M. Cannon. 1988. Vegetable crop potential in
Louisiana. Louisiana State Univ. Agr. Ctr., Louisiana State University,
Baton Rouge.
Hudson, W.D. 1986. Relationships of
domesticated and wild Physalis
philadelphica. p. 416-432. In: W. G. D'Arcy (ed.).
Solanaceae, Biology and Systematics. Columbia Univ. Press, New York.
Laidig, G.L., E.G. Knox, and R.A. Buchanan. 1983. Underexploited crops.
p. 38-64. In: W.R. Sharp, D.A. Evans, P.V. Ammirato and Y. Yamada
(eds.). Handbook of plant cell culture, crop species. Macmillan, New
York.
Menzel, M.Y. 1951. The cytotaxonomy and
genetics of Physalis.
Proc. Amer. Phil. Soc 95:132-183.
Menzel, M.Y. 1957. Cytotaxonomic studies
of Florida coastal species of Physalis.
Yrbk Amer. Phil. Soc. 1957:262-266.
Moriconi, D.N., H. Flores, and M.C.
Rush. 1988. Somatic embryogenesis and plant regeneration in Physalis ixocarpa.
Brot. Plant Physiol. Suppl. 86:118. (Abstr.)
Mosino-Aleman, P.A., and E. Garcia. 1974. The climate of Mexico. p.
345-404. In: R.A. Bryson and F. K. Hare (eds.) World survey of
climatology Vol. 11. Climates of North America. Elsevier Scientific
Publ. Co. New York.
Mulato-Brito, J., L.
Jankiewicz, V.M. Fernandez-Orduna, F. Cartujano-Escobar, and L.M.
Serrano-Covarrubias. 1985. Growth, fructification and plastochron index
of different branches in the crown of the husk tomato (Physalis ixocarpa
Brot.). Acta Soc. Bot. Pol. 54:195-206.
Murashige, T., and F. Skoog. 1962. A revised medium for rapid growth
and bioassays with tobacco tissue cultures. Physiol. Plant. 15:473-497.
National Climatic Data Center. 1985. Climatography of the United States
No 20. Climate summaries for selected sites, 1951-80. Louisiana.
Depart. of Commerce, Asheville, NC.
Pandey, K.K. 1957. Genetics of
self-incompatibility in Physalis
ixocarpa Brot.: a new system. Amer. J. Bot. 44:879-887.
Quiros, C.F. 1984. Overview of the
genetics and breeding of husk tomato. HortScience 19:872-874.
Raja-Rao, K.G. 1979. Morphology of the
pachytene chromosomes of tomatillo (Physalis
ixocarpa Brot.). Indian Bot. 2:209-213.
Raja-Rao, K.G., and Y. Lydia-Prasad.
1984. Pachytene chromosomes of Physalis
lanceifolia Ness. Cytologia 49:567-572.
Rick, C.M. 1987. Genetic resources in Lycopersicon. p. 17-26. In: D.J.
Nevins, and R.A. Jones (eds.). Tomato biotechnology. Alan R. Liss, New
York.
Saray-Meza, C.R., A Palacios A., and E.
Villanueva. 1978. Rendidora, nueva varieded de tomate de cascara. El
Campo 54:17-21.
Sullivan, J.R. 1984. Pollination biology
of Physalis viscosa
var. cinerascens (solanaceae). Amer. J. Bot. 71:815-820.
Sullivan, J.R. 1986. Reproductive
biology of Physalis
viscosa. p. 274-283. In: W.G. D'Arcy (ed.). Solanaceae,
biology and systematics. Columbia Univ. Press, New York.
Venkateswarlu, J., and K.G. Raja-Rao.
1977. Morphology of the pachytene chromosomes of Physalis philadelphica
Lam. Caryologia 30:435440.
Venkateswarlu, J., and K.G. Raja-Rao.
1979a. Morphology of the pachytene chromosomes of Physalis pubescens
L. Cytologia 44:161-166.
Venkateswarlu, J., and K. G. Raja-Rao.
1979b. Morphology of the pachytene chromosomes of Physalis angulate
L. Cytologia 44:557-560.
Vietmeyer, N.D. 1986. Lesser-know plants
of potential use in agriculture and forestry Science 232:1379-1384.
Villanueva, E., and J. Loya-Ramirez. 1976. El cultivo de tomate de
cascara en el estado de Morelos. Circular CIAMEC N 57 Zacatepec,
Morelo, Mexico. p. 11.
Waterfall, U.T. 1958. A taxonomic study
of the genus Physalis
in North America north of Mexico. Rhodora 60:107-114.
Waterfall, U.T. 1967. Physalis in Mexico,
Central America, and the West Indies. Rhodora 69:82-120.
Willis, J.C. 1966. A dictionary of the
flowering plants and ferns. Cambridge Univ. Press, Cambridge, UK.
Yamaguchi, M. 1983. World vegetables,
principles, production and nutritive values. AVI., Westport, CT.
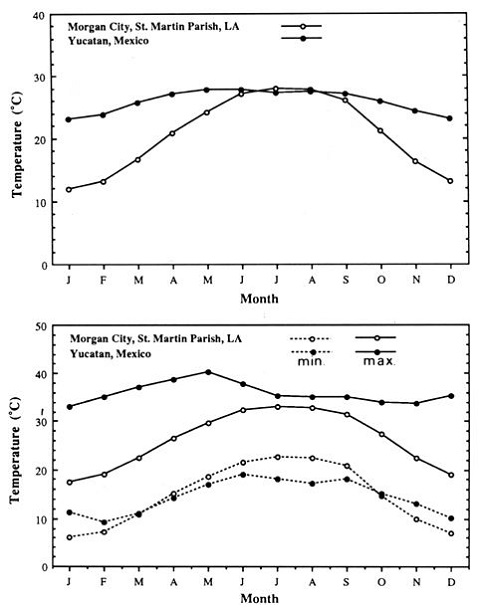
Fig. 1. Daily
mean temperatures (top), and maximum and minimum temperatures (bottom)
of Yucatan, Mexico (period 1931-1960) and Morgan City, Louisiana
(period 1951-1980). Sources: National Climatic Data Center (1985), and
Mosino and Garcia (1974).
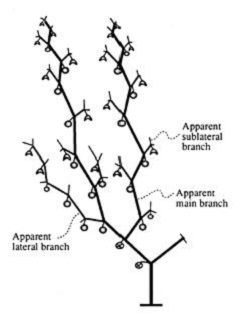
Fig. 2. Diagram
of the tomatillo plant in full development showing fruit setting over
different branches. Modified from Cartujano-Escobar et al. (1985a).
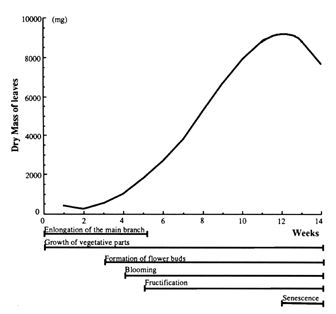
Fig. 3. Growth
curve and phenological stages of tomatillo in Morelos Mexico. Modified
from Mulato-Brito et al. (1985) and Cartujano-Escobar et al. (1985a).

Fig. 4. Tomatillo
fruit at maturity. The fruit break open the enveloping husk. The fruit
are greenish-yellow, with a slightly sticky surface, high in solids and
containing many seeds.
Last update September 4, 1997 by aw
|
|